
Swimming Behaviour of the Ciliate Paramecium in Weightlessness
(TEXUS 27 and TEXUS 28)
R. Hemmersbach
Institute of Aerospace Medicine, DLR
(German Aerospace Research Establishment), D-51140 Köln,
Germany
Six minutes of free-fall conditions during the parabolic
flights of the TEXUS 27-28 sounding rockets were sufficient to
induce behavioural changes in Paramecium. The centrifugal spin
during launch was perceived by Paramecium, resulting in a higher
precision of the negative gravitaxis than in 1 g. After onset of
microgravity, paramecia maintained their former swimming
direction for about 30 s. The ensuing random swimming indicated
that gravity was indeed the stimulus for the observed orientation
at 1 g. The individual swimming tracks remained straight in the
absence of gravity, indicating that no depolarisation of the
membrane to the level of an action potential had occurred. The
mean swimming velocity increased significantly, indicating
hyperpolarisation of the membrane and/or changes in the amount
of cAMP. The increase in velocity was transient and subsided
after 3 min to the speed of the former horizontal swimming at 1
g. Electron microscopy revealed that trichocysts, organelles
which may be extruded through mechanical stress, were still
present within Paramecium after the experiment.
Introduction
How is the gravity stimulus perceived on the cellular level?
Gravitactic protozoa offer the advantage in studying how the
physical signal of gravity is transformed into a physiological
response. The freshwater ciliate Paramecium uses gravity
as an environmental stimulus for its spatial orientation
(gravitaxis) and for the control of its swimming velocity
(gravikinesis). Though Paramecium is heavier than water,
it is able to swim against the direction of gravity (negative
gravitaxis). Measurement of the swimming velocities in the
different swimming directions revealed that paramecia antagonise
sedimentation by an active speed regulation, resulting in a
faster upward swimming velocity than expected by
calculations.1,2 In the absence of other stimuli,
gravitaxis and gravikinesis help the cells to move and to stay
in suitable habitats.
The phenomena of the graviresponses are well described;
however, the basic mechanisms are still under discussion. Drawing
on our current knowledge of the perception of other environmental
stimuli such as light³ and chemicals,4 a
physiological signal transduction chain can be postulated for
gravitaxis and gravikinesis. Cellular elements are discussed as
possible gravireceptors in Paramecium: the whole cytoplasm
via opening of specific ion channels in the membrane of the cell
or heavy cell organelles in interaction with the cytoskeleton and
membranes.2, 5
Paramecium offers the advantage that changes in
membrane potential and/or second messengers can be identified by
corresponding changes in the swimming behaviour.6, 7
Hyperpolarisation and/or an increase in cAMP augments the ciliary
beat, thus raising the forward swimming velocity. On the other
hand, depolarisation to the level of an action potential and/or
an increase in cGMP induces a reversal of the ciliary beat
direction and thus backward swimming.
An organism such as Paramecium with clear
graviresponses might change its behaviour after transition to
weightlessness, thus providing insight into the mechanism of
graviperception.
Materials and methods
Paramecium biaurelia in their original culture medium
(straw medium, pH 7.2) were enriched without centrifuging to
exclude acceleration effects before the beginning of the
experiment. Cells were placed in a temperature-controlled (via
Peltier elements) observation chamber (diameter 22 mm; depth 0.5
mm) consisting of two titanium frames housing two pieces of glass
(RG 645, Schott & Gen., Mainz, Germany). The sample was placed
45 min before launch on the stage of the horizontal microscope
within the rocket's module (TEM 06-19). The swimming cells were
observed in dark field mode with a 1.6x objective at 22° C.
The cells were again observed 45 min after landing in a
horizontal microscope and afterwards washed in EGTA (0.5 mM),
fixed in 1% OsO4(0°C) for 30 min and further
prepared for electron microscopy.8
Accelerations
A mean linear acceleration
of 5 g and a centrifugal (spin) acceleration acted during the
first 50 s of the rocket's boost phase. Only the spin
acceleration, peaking at 6.4 g, remained just before the onset
of µg, and was stopped within 1 s. When the video signal was
received 58 s after lift-off, the residual acceleration had
dropped to 2.6x10-4 g and declined to
<4x10-6g at the sample's position.
Vibration test
Cells were exposed to a
vibration test (5.5 g, 20-2000 Hz for 1 min) simulating the
worst-case Skylark rocket launch (MBB-ERNO, Bremen, Germany).
Slow rotating centrifuge microscope
Hypergravity experiments (up to 5 g) were performed on the
Niedergeschwindigkeits-Zentrifugenmikroskop (NIZEMI) rotating
centrifuge (Dornier, Friedrichshafen, Germany). The optical
conditions were as in the flight experiment.9
Fast-rotating clinostat
A fast-rotating
clinostat microscope, developed by Briegleb and co-workers, was
used to cancel the influence of gravity by reducing the
presentation time of the gravitational stimulus.9, 10
At 60 rpm, the centrifugal force amounts to 0 g in the centre and
to 4x10-2g at the border of an observation chamber of
15 mm radius.
Video analysis
Evaluation was performed
by computer-controlled realtime online image analysis as well as
using video-recorded sequences.11, 12 Data for cell
orientation, swimming velocities and linearity of individual
swimming tracks were obtained. The degree of orientation was
quantified by the Rayleigh test13 which yields a
statistical value (r-value) that runs from 0 (random) to 1
(perfect orientation of all organisms in the same direction).
Results
For simulating the effect of linear acceleration and
vibrations during rocket launch, negative gravitactic cultures
of Paramecium were exposed to 5 g and a vibration profile
for 1 min. Gravitactic behaviour was maintained in both cases.
Reorientation under 5 g towards the resulting acceleration vector
occurred within seconds. Orientation precision was higher than
under 1 g. Exposure to hypergravity decreased the mean swimming
velocity, while vibration had no effect. The relationship between
the up, down and horizontal velocities remained
unaffected.14, 9
Before lift-off, Paramecium showed a precise negative
gravitaxis. Most of the cells swam upwards with a high precision
in orientation (r-value = 0.55; theta = 9°) (Fig. 1a & 1d;
Fig. 2). After 1 s of free-fall, when the video signal was
received from the rocket, paramecia still had a residual
orientation, but towards 90±30° (theta = 96.2°),
that is, towards the centre of the rocket (Fig. 1b & 1e; Fig. 2).
The degree of orientation was even higher (r-value = 0.89) in
comparison to the behaviour at 1 g. Random swimming was
registered after about 80 s in µg (r-value: 0.01) (Fig. 1c
& 1f; Fig. 2). Paramecium again showed negative gravitaxis
immediately after payload retrieval.9
During the whole µg period, the linearity of the
individual swimming tracks was the same as under 1 g. Swimming
direction reversals and cell movement cessation were not
observed.9
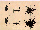
Fig. 1. Representative circular histograms from TEXUS 27 (a-c)
and TEXUS 28 (d-f) showing the swimming directions of Paramecium
at different times: negative gravitaxis 5 min before lift-off (a,
d); residual orientation 1 s after µg onset towards the
centre of the rocket (originally induced by the centrifugal
launch accelerations) (b, e); random distribution 80 s after
µg onset. Each histogram represents >300 tracks.
The cells' mean swimming velocity measured during the total
free-fall period was higher (4.3% TEXUS 28; 7.5% TEXUS 27), than
under 1 g. However, the increase was transient: after 3 min in
µg the mean swimming velocity approached the value of the
former horizontal swimming velocity (Fig. 2). The differences
between the upward, downward and horizontal swimming at 1 g
before launch disappeared under free-fall conditions, but were
re-established after landing.14
Experiments on a fast-rotating clinostat on the ground
revealed similar changes in orientation behaviour, albeit with
some temporal delay. After starting rotation, a residual
orientation in the former upward direction was measured for about
120 s, followed by random distribution. Paramecium
regained its negative gravitaxis in the first minute after
stopping the clinostat, even after a 2 hr run. During the run,
Paramecium exhibited an increased swimming velocity as in
the rocket. However, the increase was not transient but
stable.9, 14

Fig. 2. Degree of orientation (r-value) (bars) and
corresponding mean swimming velocities (line) before launch (1
g), during µg (0 g) and after landing (1 g). Measurements
were performed continuously at 20 s intervals (N = 5809).
Trichocysts in Paramecium were taken as a stress
indicator during the experimental conditions. Exocytosis of
trichocysts can be provoked, for example, by an increase in the
permeability of the membrane to calcium, by shear forces or by
cell damage.15 Paramecia exposed to sounding rocket
flights with periods of accelerations contained trichocysts in
their normal condensed morphology, still attached to the cellular
membrane.8
Discussion
Experiments under the conditions of brief weightlessness do
provide sufficient time to observe behavioural changes of
gravitactic unicellular organisms and to provide further clues
on the nature of the receptor mechanism.
Random distribution of the paramecia in µg proved that
gravity is indeed the stimulus for the observed orientation at
1 g. Before a random distribution of the cells was measured,
paramecia showed a residual orientation, induced by the last
effective acceleration. Results obtained on the NIZEMI slow-
rotating centrifuge support the conclusion that the first
observed swimming direction in the rocket was originally induced
by the centrifugal spin acceleration during launch. The knowledge
of this residual effect is important for the design of further
experiments.
During the 6 min free-fall conditions of the TEXUS experiment,
Paramecium showed a transient increase in the mean
swimming velocity. As supported by simulation experiments, this
is presumably not induced by acceleration or vibration during
launch. The fact that gravikinesis was not switched off
immediately indicates a slow relaxation time constant of the
gravikinetic response. The temporal delay between the gravitactic
and the gravikinetic response rekindles the question whether both
graviresponses are controlled by the same mechanism. Concerning
the mechanism of graviperception in Paramecium, it is
proposed that the whole cytoplasm acts as a statolith by exerting
a pressure of about 0.1 Pa on the lower cell
membrane,2 thus initiating a signal transduction
cascade by stimulation of ion channels in the membrane. Suitable
candidates as primary gravireceptors might also be heavy cell
organelles in interaction with the cytoskeleton and/or membranes
as in plant systems.16, 17 Further studies using
inhibitors and mutants might prove this hypothesis. Experiments
on clinostats and centrifuges are necessary for the explanation
and statistical security of data obtained from experiments under
the conditions of real µg.
An increasing number of experiments have been performed in
recent years on sounding rockets dealing with questions from
gravitational biology. Fast biological responses are well suited
for investigation aboard this facility. Late access and early
retrieval make this programme attractive for biological
experiments. MAXUS, providing 12 min of free-fall, offers further
possibilities. A centrifuge microscope on MAXUS 2 will allow us
to determine the threshold for graviperception in different
unicellular specimens.
Acknowledgements
The authors would like to thank D. Franke and his TEXUS-team
of MBB-ERNO, Bremen, Germany, and G. Schmitt and his team from
Kayser-Threde, Munich, Germany, for their excellent work. Thanks
are also expressed to the German Minister for Research and
Technology (BMFT), the German space agency (DARA) and the
European Space Agency (ESA) for support.
References
-
Hemmersbach-Krause, R., Briegleb, W. & Häder, D.-P. (1991).
Dependence of gravitaxis in Paramecium on oxygen. Eur.
J. Protistol. 27, 278-282.
-
Machemer, H. & Bräucker, R. (1992). Graviperception and
graviresponses in ciliates. Acta Protozool. 31, 185-214.
-
Colombetti, G. (1990). New trends in photobiology: photomotile
responses in ciliated protozoa. J. Photochem. Photobiol.
B4, 243-259.
-
Van Houten, J. (1992). Chemosensory transduction in eukaryotic
microorganisms.Annual Rev. Physiol. 54, 639-663.
-
Bean, B. (1984). Microbial geotaxis. In Membranes and Sensory
Transduction. (eds. Colombetti, G. & Lenci, F.). 163-198.
Plenum Press, New York.
-
Naitoh, Y. & Eckert, R. (1969). Ionic mechanisms controlling
behavioral responses of Paramecium to mechanical
stimulation. Science 164, 963-965.
-
Preston, R. R. & Saimi, Y. (1990). Calcium ions and regulation
of motility in Paramecium. In Ciliary and Flagellar
Membranes. (ed. Bloodgood, R. A.). 173-194. New York, Plenum
Press.
-
Hemmersbach-Krause, R., Briegleb, W., Häder, D.-P. &
Plattner, H. (1991). Gravity effects on Paramecium cells:
An analysis of a possible sensory function of trichocysts and of
simulated weightlessness on trichocysts exocytosis. Eur. J.
Protistol. 27, 85-92.
-
Hemmersbach-Krause, R., Briegleb, W., Häder, D.-P., Vogel,
K., Grothe, D. & Meyer, I. (1993). Orientation of Paramecium
under the conditions of weightlessness. J. Euk.
Microbiol. 40 (4), 439-446.
-
Briegleb, W. (1988). Ground-borne methods in gravitational cell
biology. The Physiologist 31, 44-47.
-
Häder, D.-P. & Lebert, M. (1985). Real time computer-
controlled tracking of motile microorganisms. Photochem.
Photobiol. 42, 509-514.
-
Häder, D.-P. & Vogel, K. (1991). Simultaneous tracking of
flagellates in real time by image analysis. J. Math. Biol.
30, 63-72.
-
Batschelet, E. (1965). Statistical methods for the analysis of
problems in animal orientation and certain biological rhythms.
In Animal Orientation and Navigation (eds. Galles, S. R,
Schmidt-Koenig, K., Jacobs, G. I & Belleville, R. ). 61-91. NASA,
Washington DC.
-
Hemmersbach-Krause, R., Briegleb, W., Vogel, K. & Häder, D.-
P. (1993). Swimming velocity of Paramecium under the
condition of weightlessness. Acta Protozoologica, 32, 229-
236.
-
Plattner, H. (1987). Synchronous exocytosis in Paramecium
cells. In Cell Fusion (ed. Sowers, A. E.). 69-98.
Plenum Press, New York.
-
Sack, F.D. (1991). Plant gravity sensing. Int. Rev.
Cytol. 127, 193-252.
-
Sievers, A., Buchen, B., Volkmann, D. & Heijnowicz, Z. (1991).
Role of the cytoskeleton in gravity perception. In The
Cytoskeletal Basis of Plant Growth and Form (ed. Lloyd, C.
W.). 169-182. Academic Press, London.
About|
Search|
Feedback



 SP1206
SP1206
Published April 1997.
Developed by ESA-ESRIN ID/D.

